
Basic Botany
Paula Dinius, Urban Horticulturist, Chelan County Extension, Washington State University
The simple plants (opens in new window) such as algae, and microorganisms (bacteria, fungi, etc.), are of great economic importance to us in both positive and negative ways. Oceanic and other aquatic algae produce a tremendous amount of oxygen while removing carbon dioxide from the atmosphere. Fungi are important for the many fermentation processes which give us our wines, cheeses, antibiotics, and other economically important products, in addition to the decomposition of past living things. On the negative side there are species of both bacteria and fungi that can cause diseases, injury, or death—of both plants and animals.
Distinguishing Characteristics of Plants
Table 1. Some distinguishing characteristics of plants.
| Kingdom | Plants | |||||
|---|---|---|---|---|---|---|
| Tracheophytes (vascular, “higher” plants) | Bryophytes (non-vascular plants) | |||||
| Phylum/Division | Spermatophyta (seed plants) | Pteridophyta (ferns) | Mosses | Liverworts | ||
| Subdivision | Magnoliophyta Angiosperms: covered seeds | Coniferophyta Gymnosperms: naked seeds | No Data | No Data | No Data | |
| Reproduction | Reproduce by seeds | Reproduce by spores | Fused gametes | Fused gametes | ||
| Classes | Liliopsidae Monocots | Magnoliopsidae Dicots | Pinopsida Polycots | multiple | multiple | multiple |
| Cotyledons (if present) | One seed leaf | Two seed leaves | Multiple seed leaves | No Data | No Data | No Data |
| Leaf shape or venation | Parallel veined leaves | Branched or net-veined leaves | Needles or scales | Leaf-like structures | No true leaves; single cell layer | Lobe-shaped |
| Flowers or flower parts | Flower parts in 3s or multiples of 3 | Flower parts in multiples of 4 or 5 | No flowers (modified bracts) | No Data | No Data | No Data |
| Vascular pattern | Xylem and phloem paired in “vascular bundles” | Xylem and phloem in cambium layer around the stem | Xylem and phloem in cambium layer around the stem | Vascular strands in diverse patterns | No Data | No Data |
| Specialized growth tissue | No cambium layer | Cambium layer | Cambium layer | No Data | No Data | No Data |
| Primary growth regions | Growth from base of plant and/or a “primary thickening meristem” | Growth at root and shoot tips plus cambium layer | Growth at root and shoot tips plus cambium layer | No Data | No Data | No Data |
| Examples | Grasses, palms, lilies | Broadleaf plants, maple, rose | Conifers, arborvitae, pine | Ferns | Sphagnum | Calyptogeia |
Table 1 describes how plants are divided. They are divided into two broad groups: Tracheophytes, which are vascular or “higher” plants, and Bryophytes, which are non-vascular plants. Tracheophytes include seed plants and ferns. Among the seed plants, angiosperms (such as flowering plants) produce covered seeds, while gymnosperms
(such as conifers) produce naked seeds. Ferns, on the other hand, reproduce by spores instead of seeds. Bryophytes include mosses and liverworts, both of which reproduce by fusing gametes rather than by seeds or spores.
The classification of plants can be described by comparing several major groups. Liliopsidae Monocots have one seed leaf (cotyledon). Their leaves are parallel-veined, and their flowers appear in parts of three or multiples of three. Their vascular system has xylem and phloem paired in vascular bundles, with no cambium layer present. Growth occurs from the base of the plant or from a primary thickening meristem. Examples include grasses, palms, and lilies.
Magnoliopsidae Dicots possess two seed leaves. Their leaves are branched or net-veined, and their flowers occur in multiples of four or five. The vascular system consists of xylem and phloem arranged in a cambium layer around the stem. They have a cambium layer for specialized growth and grow at both root and shoot tips, as well as in the cambium layer. Examples are broadleaf plants such as maple and rose.
Pinopsida Polycots have multiple seed leaves. Their leaves are needle-like or scale-like, and they do not produce flowers but instead have modified bracts. Their vascular system features xylem and phloem in a cambium layer around the stem, with a cambium layer present for growth. Growth occurs at root and shoot tips plus the cambium layer. Examples include conifers such as arborvitae and pine.
Another group of plants, represented here simply as multiple categories, have diverse traits. Ferns have leaf-like structures rather than true leaves, with vascular strands in varied patterns, and they reproduce without flowers. Sphagnum mosses do not have true leaves but instead have a single cell layer. They also lack vascular tissues as seen in higher plants. Calypogeia, a type of liverwort, has lobe-shaped leaves.
Plant Taxonomy
Plant taxonomy is not an exact science. There are a few different schemes, and these change when new and better information is available. The most recent information providing taxonomic data can be accessed through an organization called the Interagency Taxonomic Information System (ITIS).
It was formed through partnership of many federal agencies in 2000. ITIS uses data standards from the International Code of Botanical Nomenclature, the International Code of Nomenclature of Bacteria, and ICN of Zoology; and it uses the Seven Kingdom system: Bacteria, Protozoa, Plantae, Fungi, Animalia, Chromists, and Archaea.
Gardeners, of course, will primarily concern themselves with the phyla/divisions of the plant kingdom. In the plant kingdom the ranking of division (which is the equivalent of phylum in other kingdoms) is ranked below kingdom and above class. But gardeners will learn about other kingdoms and phyla when they study pests and beneficial organisms, including insects, diseases, and microbes in the soil food web.
As practical horticulturists, we are primarily concerned with higher plants (vascular plants), of which the two most important groups are the subdivision Magnoliophyta, or angiosperms (opens in new window) (flowering plants with covered seed), and Coniferophyta, or gymnosperms (opens in new window) (naked-seed plants, such as conifers). Also of some importance in the vascular plant group are the ferns, which belong to the division Pteridophyta (spore-bearing plants).
The Magnoliophyta subdivision is split into two classes: the Magnoliopsida (broadleaf plants with two cotyledons (opens in new window), casually known as dicotyledons (opens in new window) or “dicots”) and the Liliopsida (plants with one cotyledon: grasses, palms, and lilies, casually known as monocotyledons or “monocots”).
Seed plants are those vascular plants which reproduce by seeds. Gymnosperms produce seed in a cone or cone-like structure, and the seed does not have a covering. Plants which are in this category are in the class Pinopsida, the conifers, and include pines, firs, spruces, hemlocks, cedars, arborvitae, and others.
Angiosperms produce seed which have some sort of a covering, such as an apricot (Prunus armeniaca) that has a “pit,” or the tiny seed inside the woody capsule of a rhododendron fruit. Angiosperms are divided into two very broad groups: monocots and dicots. Seeds of monocot plants, when germinating, have one cotyledon, or seed leaf. Another characteristic of the monocots is that the veins in each leaf run parallel to the mid-rib of the leaf. Monocots include grasses, palms, and lilies.
Germinating seeds of dicot plants display two cotyledon leaves. The seed leaves of dicots usually do not look anything like the true leaves which emerge later. The seed leaves contain the stored food of the seed which is used for the initial growth of the seedling. Leaves of dicots have reticulate (opens in new window) or net veining instead of the parallel veining characteristic of the monocots.
Ferns are plants that have root-like structures called rhizomes (opens in new window) and leaf-like structures called fronds (opens in new window). Fern reproduction is by means of spores, not seeds, which are produced in sacs on the underside of the fronds.
Table 2 shows examples of taxonomic rankings of three plants using the additional ranks of variety and cultivar (cultivated variety) as outlined in the botanical code.
Flowers and fruits are the reproductive parts of a plant—they make or contain seeds. Vegetative material is the rest of the plant. Botanically speaking then, tomatoes, corn, and squashes are fruits, not vegetables, because they contain seeds. Onions, celery, and lettuce are vegetables in both the botanical and culinary senses.
Plant Classification According to Life Cycle
Plants may also be classified as to whether they are annual, biennial, or perennial. Knowing the life cycle of plants is not only important for planting plants but also for weed control.
Controlling an annual weed may take entirely different measures than controlling a perennial weed.
Table 2. Examples of taxonomic classification: pear, spruce, and cauliflower.| Classification Unit | Aristocrat Pear | Colorado Blue Spruce | Cauliflower |
|---|---|---|---|
| Kingdom | Plantae | Plantae | Plantae |
| Division | Magnoliophyta | Coniferophyta | Magnoliophyta |
| Class | Dicotyledons | Pinopsida | Dicotyledons |
| Order | Rosales | Pinales | Brassicales |
| Family | Rosaceae | Pinaceae | Brassicaceae |
| Genus | Pyrus | Picea | Brassica |
| Species | calleryana | pungens | oleracea |
| Botanical Variety | No Data | glauca1 | botrytis2 |
| Cultivated Variety or Cultivar | ‘Aristocrat’3 | ‘Montgomery’4 | ‘Snow Crown’5 |
Note: The genus and species name plus any botanical variety or cultivated variety name make up the scientific name or botanical name of a plant.
For more on genus, species, cultivated varieties, and naming of plants, see Chapter 2: Plant Nomenclature and Identification.
Source: PLANTS Database.
1 The naturally occurring glauca (blue needled) Colorado spruce is a form of the normally green-leafed Colorado spruce.
2 Compare to varietal name Brassica oleracea capitata which is cabbage, while Brassica oleracea gemmifera is Brussels sprouts.
3 A selected upright form of the Callery pear.
4 A selected slow growing, bushy form of the Colorado blue spruce.
5 White, self-blanching, drought-resistant All-American Selection Winner.
Annuals (opens in new window) are plants which complete their entire life cycle—germinate, mature, and set seed—in one growing season. Summer annuals start growing in the spring of the year, grow, flower, go to seed, and die by mid summer to fall. Winter annuals generally germinate during the late summer to fall, flower from fall to spring, and then, depending on the species, set seed and die. Hardy annuals have a certain degree of frost tolerance.
Biennial plants (opens in new window) complete their life cycle in two physiological growing seasons, which is normally two years. The first year the plants grow vegetatively, usually as a rosette (opens in new window) close to the ground. The chill of the winter season causes a biennial plant to set up flower buds. This chill-to-reproduce process is called vernalization (opens in new window). During the following growing season, the plant will flower, set seed, and die. The biennial process requires a warm period for vegetative growth followed by a cool period for flower bud formation, followed by another warm period for flower and seed development. The two physiological growing seasons are two warm periods interrupted by a chill period. This can be, and unfortunately often is, accomplished in less than a year, in a process known as “bolting.”
One example of biennial plants is the cabbage family: Brassica. If not harvested but left in the garden to overwinter and grow the second year, brassicas will produce nice yellow flowers (and seeds). However, to get a jump on the season, many times kale, cabbage, kohlrabi, etc. will be started early (Jan., Feb., Mar.) in greenhouses where the plants will grow vegetatively. The plants may then be set out as early as March, April, or May. But if cold temperatures occur (40°F or colder), after plants are planted outdoors, they may think they have gone through a winter and set up flower buds and bloom instead of producing their typical head or bulb.
Perennial plants (opens in new window) live for three or more years. Herbaceous perennials (opens in new window) have stems which die back each winter. Woody perennials (opens in new window) have aboveground stems which will live through the winter. Shrubs are woody plants that have basal activity (can sprout new branches from the base of the plant). Trees are considered to be single- to few-trunked woody plants.
Tender perennials have limited frost tolerance. See the Plant Hardiness section later in this chapter for further discussion of plant survival.
A number of tender perennial bedding and vegetable plants are used as annuals in our climate. Examples include wax begonia (Begonia semperflorens), Coleus sp., and tomato (Solanum lycopersicum).
Plant Anatomy
Plants are made up of cells which are grouped into tissues, based on similar characteristics. These tissues make up the structures, or organs, of a plant: roots, stems, leaves, flowers, fruit, and seeds. Each of these organs has been developed for a specific purpose or task.
Plant Cells
The basic building unit of the plant is the cell (Figure 1). There are many different types of plant cells. The size, shape, and internal contents of each cell will vary according to the function of the individual cell.
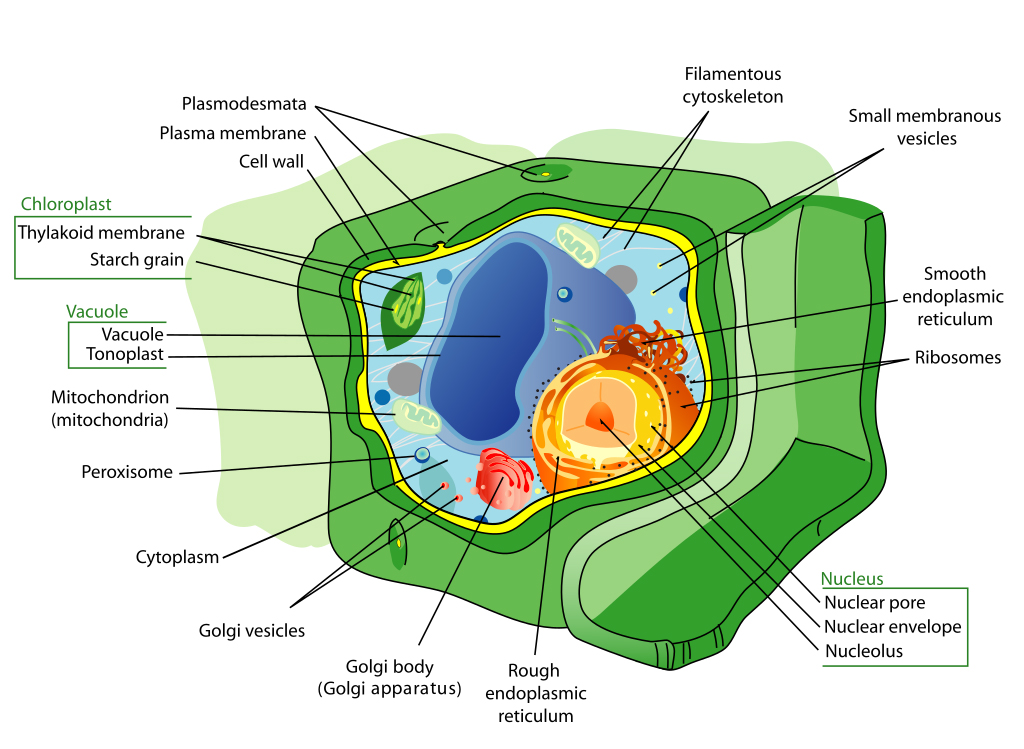
The “typical” plant cell is surrounded by a cell wall that keeps the internal contents contained. Thin-walled cells are usually flexible, such as those one would find in leaves and flower petals. Cell walls may also be thicker and more rigid, such as those found in the cells of woody plant stems or walnut husks. Inside the cell wall is a semi-permeable membrane which allows water and dissolved materials to enter and exit the cell.
The internal living material within a cell is called protoplasm (opens in new window). The protoplasm may have a number of different constituents depending, again, on the particular function of that cell in the total activity of the plant. The cytoplasm is everything within the protoplasm minus the nucleus. Cytoplasm (opens in new window) includes organelles suspended in a gelatinous material which moves around in the cell via a process called cytoplasmic streaming (opens in new window).
The nucleus could be called the brains of the cell, because it contains all the genetic information to form new cells and, in most cases, an entire new plant. Another cell component, the vacuole, is a membrane-bound organelle which regulates cell turgor pressure (rigidity or flaccidity of the cell) and acidity. It also stores and transports proteins and carbohydrates, along with waste products and other byproducts of metabolism (opens in new window).
Organelles are smaller structures that have specific functions within a cell. For instance, mitochondria (opens in new window) are the powerhouses of cells, controlling the various chemical functions (metabolism) of a cell. Another important organelle is the chloroplast (opens in new window), which contains the chlorophyll (opens in new window), a green pigment that absorbs light to provide the energy for photosynthesis.
It is important to stress that cells of different kinds vary in size, shape, and content, depending on the cell’s location and function within the plant. Cells in the various plant tissues and from different parts of the plant are extremely variable.
Cell Division. Cells in certain regions of a plant divide by a process called mitosis (opens in new window). A cell can divide to form two identical daughter cells; these two daughter cells may then divide, and so on. This cell division happens in specific areas called meristems (opens in new window), or meristematic zones. There are two primary, or apical meristems (opens in new window), one at the tip of each growing shoot, and the other at the tip of each growing root. A secondary meristem increases the diameter of woody plant stems and is called the cambium (opens in new window).
Plant Tissues and Organs
Plant tissues are groups of cells which are alike in structure and function. Types of plant tissues include the epidermis (opens in new window), meristem (opens in new window), parenchyma, sclerenchyma, xylem (opens in new window), phloem (opens in new window), and cambium (opens in new window).
Epidermis. This is the outermost layer of a plant, the “skin” if you will. This single layer usually has a waxy covering, called cutin, to reduce water loss.
Meristem. Meristem tissues are made of actively dividing cells. Meristems are the areas from which active growth springs.
Parenchyma. Parenchyma tissue has thin-walled, simple cells. Parenchyma tissue in leaves is responsible for photosynthesis. Parenchyma cells can become meristematic in order to grow over a wound.
Sclerenchyma. The cells of sclerenchyma tissue have thick walls to support a plant.
Xylem. This structurally complex tissue has cells that interlock and form tubes to conduct water and nutrients from roots to the rest of the plant. As the cells and tissues mature, they form the wood within woody plants.
Phloem. This tissue is also made of pipe-like cells, but they conduct food and the products of photosynthesis throughout a plant, including back to the roots.
Cambium. This is a layer of active growth and development of xylem and phloem.
When multiple tissues are put together into a much larger unit for a specific function, they form a plant organ. The different plant organs (stems, roots, leaves, and flowers) are discussed in sections below.
Stems
Stems on vascular plants are varied as to their function, structure, and form. Stems grow by increasing in length, and also by increasing in diameter, in the case of woody stems.
Stem Function. Stems have numerous functions in a plant, including support, transport of water, nutrients, and other compounds, and storage of those materials.
Stems support and extend the plant parts, such as leaves, upwards from the roots. This extension may be minimal as in the case of some low growing grasses, low growing herbaceous plants, and ground covers. Stems may also extend the leaves well above the soil as with taller growing grasses, herbaceous plants, shrubs, and trees.
Stems transport water, nutrients, and materials from the roots into the crown (opens in new window) (all aboveground parts) of the plant. Food and other materials produced by the leaves are transported down toward the roots and also toward the end of the stem, or the stem-growing point.
Stems can store reserve nutrients over the winter to be used for the first flush of growth in the spring. Sometimes stems are modified into specialized structures for storing materials like starches and water.
Stems are said to be part of the shoot system (opens in new window) (stems and leaves) of the plant and originate in a bud (opens in new window). Cells in the apical meristem at the tip of a bud start to divide very rapidly when growth starts in the spring. Behind these dividing cells, the just-completed new cells start to elongate and push the tip further out. This is called the zone of elongation (opens in new window). Behind the zone of elongation the cells start to change and differentiate, or mature, into specialized plant tissues such as phloem, cambium, xylem, and bud and leaf initials. This area is called the zone of maturation (opens in new window), or the zone of differentiation (opens in new window).
Stem Structure. A cross-sectional view of a stem of any common plant will present evidence to show what category of plant it is: a woody plant, an herbaceous dicot, or an herbaceous monocot. Each of these categories has a different cross-sectional configuration of the vascular system.In cross section, a woody stem will display a series of concentric rings. The very outside layer is bark which, itself, is made up of a few different layers. The primary purpose of the bark is to protect the stem. It can be quite thick on some species and very thin on others. Bark is usually thinner on young woody plants and thicker on older plants. It may be smooth or furrowed. In some species it may peel off in elongated strips or patches in a process called exfoliation. Bark characteristics vary with the species and often can be diagnostic for species identification.
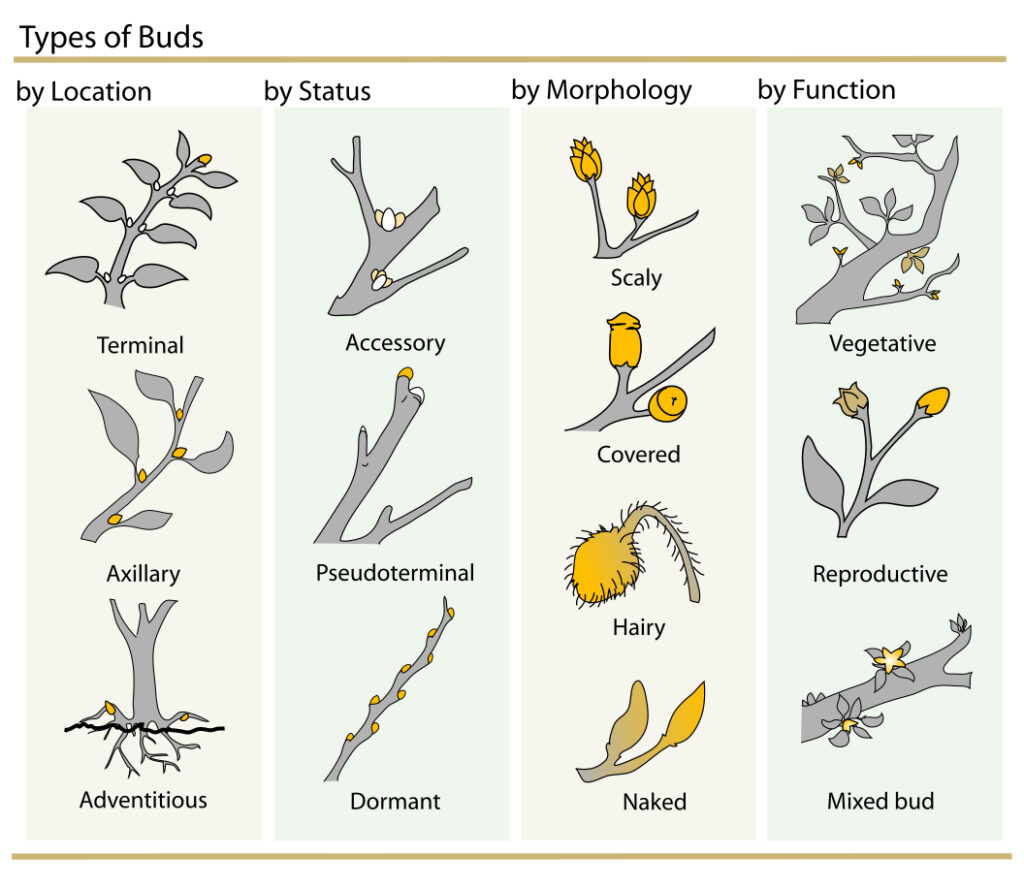
Plant Bud Terminology
Plant buds can be described in terms of where they occur and how they are arranged (location), when they will be active (status), what they look like (morphology), and what kinds of tissue they contain (function). Image credit: Plant cell structure (opens in new window).
Inside the bark is a tissue that is a few cells thick, called the phloem. Phloem translocates food and other compounds produced in the leaves both down toward the roots and also upward, toward the growing end of the stem.
The next layer is the cambium. This is where active cell division (growth) takes place. The cambium increases the diameter of a stem, forming phloem to its outside and xylem to the inside. On any pruning cut or other wound on a woody stem, the initial regrowth process is initiated by the cambium or xylem ray cells that run radially across the wood.

Xylem is found inside the cambium and typically forms the bulk of a woody stem. Xylem is a vascular tissue that moves water and mineral nutrients absorbed by the roots, plus other compounds produced by the root cells, toward the top of the plant. Ray cells are specialized xylem cells that can move water and nutrients laterally, that is, from the inner parts of the xylem outward toward the cambium and the bark, and from the bark area inward toward the center of the stem.
Another function of the xylem is to hold a woody stem, such as a shrub stem or tree trunk, upright. Xylem may also be called “wood” and could be considered the backbone of a tree.
Xylem tissue is divided into two distinct parts (Figure 2). The outer part, called sapwood (opens in new window), does the actual conduction from the roots to the top of the plant. Toward the center of the stem, inside the ring of sapwood, is the heartwood (opens in new window). Heartwood is plugged up sapwood and does not function as a translocation tissue anymore. However, it provides the structural rigidity that supports larger limbs and holds tree trunks upright. It is also the major portion of larger stems and trunks. The sapwood is often, but not always, lighter in color than the heartwood.
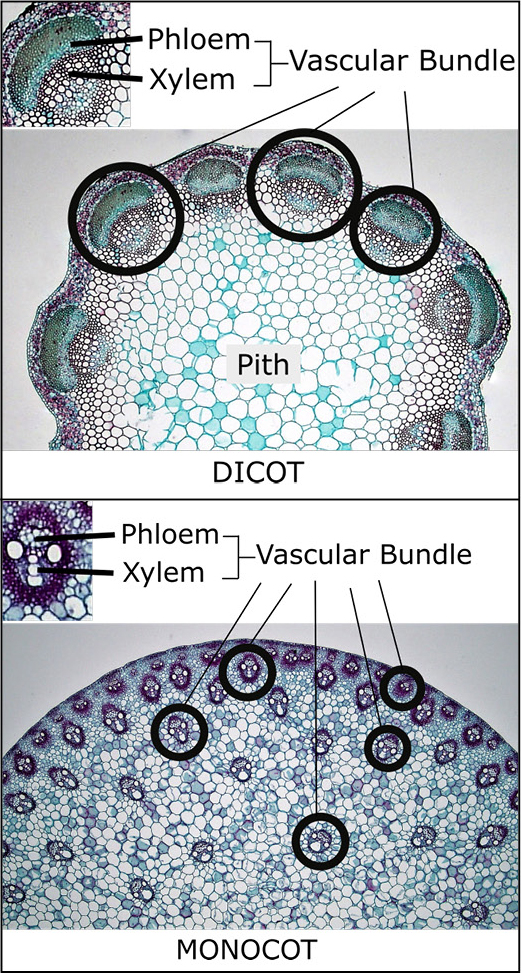
In contrast to woody dicots, herbaceous dicots have their vascular system arranged in bundles inside the stem (Figure 3). Each bundle, or vessel, has phloem on the outside, then a layer of cambium, with the xylem on the inside. A significant portion of the remaining internal part of an herbaceous dicot stem is made up of a spongy tissue called pith (opens in new window). In celery stalks (stems), the “strings” are vascular bundles of xylem and phloem. Examples include peony, kale, and salvia.
Herbaceous monocot stems also have bundled vessels, but they are scattered throughout the cross section of the stem instead of being arranged in a circular pattern. Examples of monocots are palm trees, lilies, and grasses.
Stem Morphology. The morphology of a one-year-old woody plant stem can tell a lot about how the plant grows, and many times is used in the diagnosis of plant problems.
At the tip of a stem is a terminal bud (Figure 4) which is usually covered with scales, called bud scales. Along the stem, there are either leaves or leaf scars where there once were leaves. The place where a leaf or leaves attach to a stem is called a node. The internode is the area on a stem between the nodes.
Where a leaf is attached to a stem, it forms an angle with the stem called a leaf axil. Wherever there is, or was, a leaf, there is a bud. This usually is the only place on a stem where buds are found. These are axillary or lateral buds, and they may be either simple or mixed, based on their potential. Simple buds contain either reproductive parts (flowers) or vegetative parts (leaves, shoots), but not both. Some simple flower buds may contain more than one flower bud. Mixed buds contain both reproductive and vegetative parts.
There are usually slightly raised bumps on a stem, called lenticels. The lenticels may be indistinct or quite prominent. They may appear as round or oval “blotches” on younger stems.
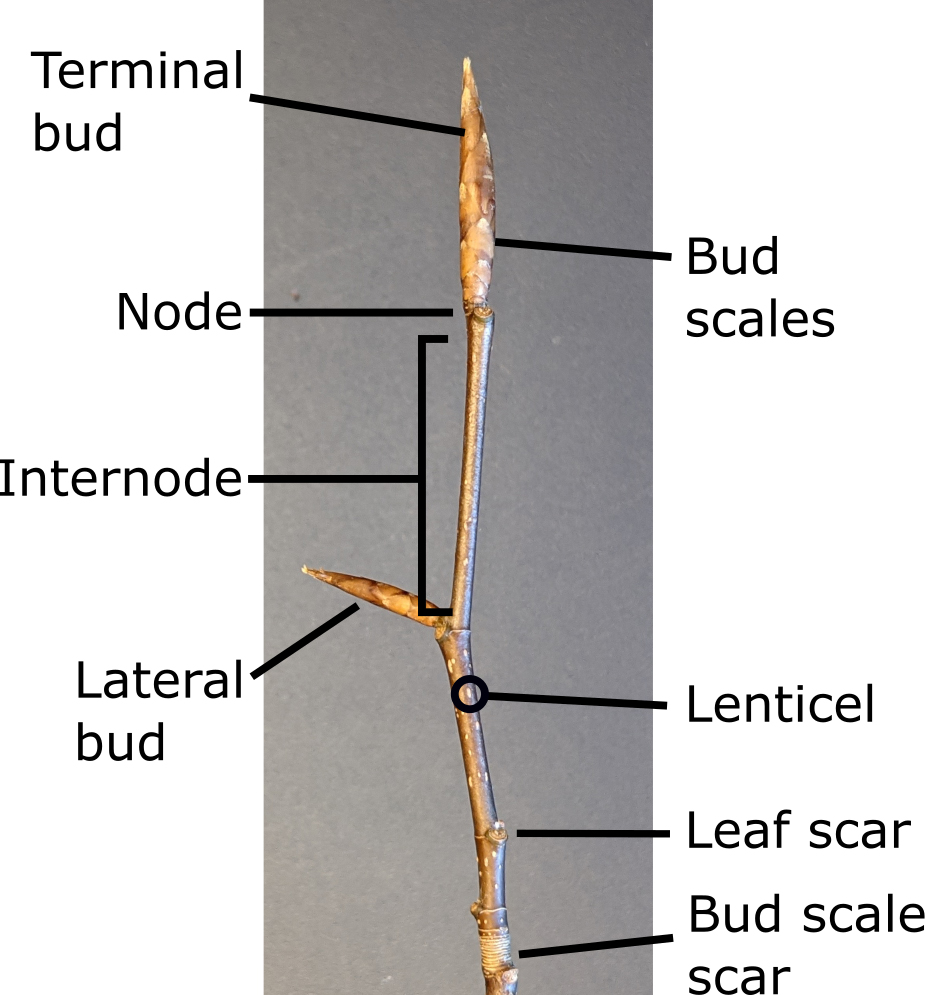
Lenticels serve as an opening for gas exchange between the outside atmosphere and the living cells within the stem. Lenticels allow oxygen from the atmosphere in and respiratory carbon dioxide from internal living cells out. Sometimes the shape, color, or other distinctive features of lenticels are helpful for plant identification.
When a plant starts to grow in the spring, the bud scales will start to enlarge and unfold away from the emerging shoot or flower. The bud scales will eventually fall off (abscise (opens in new window)), leaving a scar (a ring around the branch), called the bud scale scar (opens in new window), or scar of the terminal bud. At the end of the growing season, the distance from the newly formed terminal bud and scar where the terminal bud was at the beginning of the growing season is one season’s growth. Sometimes these scale scars may be evident for a number of years, so it may be possible to tell how well a plant was growing from year to year, based on the demonstrated length of twig growth.
If the seasonal growth is less than optimal or usual, it generally means that some sort of adverse condition affected the plant. This could be an environmental factor such as unsuitable temperature, drought, or flooding. Shorter than normal growth may also be an indication of root damage.
Specialized and Modified Stems. Stems are normally aboveground plant structures, but occasionally they will take on a different form. There are many possible modifications to the typical branch-like stem structure (Figure 5). By definition, stems should have leaves and buds.

Crowns are stems that are very short because of compressed internodes. The leaves are usually in a rosette form (many leaves emerging radially from a short stem). Examples are African violet (Saintpaulia ionanthia) and strawberry (Fragaria spp.).
Stolons (opens in new window), or runners, are elongated stems growing on top or very close to the soil surface. These runners will generally leaf out and root at the nodes. An example of this is the strawberry plant.
Rhizomes are similar to stolons, except they are underground stems and usually have parchment-like leaves at the nodes. New shoots arise at the nodes. Examples are wintergreen (Gaultheria procumbens), bunchberry (Cornus canadensis), various blue grasses (Poa spp.), and many others. There are also enlarged rhizomes, called fleshy rhizomes, which are underground food/water storage organs. Examples are bearded iris (Iris germanica), as opposed to bulbous iris (Iris reticulata), or our native iris, Douglas iris (Iris douglasiana).
Tubers (opens in new window) are enlarged underground stems, the classical example of which is the potato (Solanum tuberosum). The “eyes” are actually buds at a node, and the “skin” is actually bark. Another type of tuberous stem is that of the cyclamen (Cyclamen spp.).
Bulbs (opens in new window) are a compressed shoot surrounded by either fleshy or parchment-like leaves. Examples are onion (Allium spp.) and tulip (Tulipa spp.). An onion’s layers are modified fleshy leaves.
Corms (opens in new window) are short, enlarged underground stems used mainly for food storage. They are sometimes erroneously called bulbs. Corms contain no leaf tissue—parchment or otherwise. Examples are gladiolus (Gladiolus spp.) and crocus (Crocus spp.).
Roots
Roots are typically the underground portion of the plant usually growing in soil or some other suitable medium. Roots generally receive much less attention than the aboveground parts of the plant, as it is usually, “out of sight, out of mind.” Roots are very important to plant health, and when root problems arise, the symptoms very often are manifested in the aboveground portions of the plant; therefore, it is important to understand how roots develop in maintaining plant health.
Root Function. Roots have many functions in maintaining the health of a plant. For instance, roots anchor and stabilize a plant in a suitable growing medium or substrate, usually soil for terrestrial plants (in contrast to floating aquatic plants like duckweed).
Roots absorb water and its associated dissolved mineral nutrients from the growing medium. Dissolved mineral nutrients enter a root via the root hairs (opens in new window) (see below) by osmosis: a process whereby water and dissolved materials move across a semi-permeable membrane such as a cell membrane. The flow is from an area of greater concentration of dissolved material to an area of lower concentration of that dissolved material. There also is a lesser flow in the reverse direction.
Root Respiration. Roots absorb oxygen from the air which is found in soil pores and then give off carbon dioxide in order to convert sugars to energy used to carry out cellular work. Most roots must be in friable soil that allows the gas exchange of oxygen and carbon dioxide.
Oxygen must be available for root absorption and carbon dioxide must be able to be expelled back into the atmosphere to prevent soil carbon dioxide toxicity.
Roots produce hormones which affect various phases of a plant’s growth and development and secondary compounds for plant defense. Plant hormones, mainly cytokinins, are produced in the roots and transported upwards to stimulate cell division. They play a chemical dance with other plant hormones, most particularly auxins. More on this later in the chapter. Generally, there are two types of secondary compounds, those produced regularly in low concentrations for environmental competition and self-defense, called constitutive secondary compounds (opens in new window), and those produced in greater concentrations when triggered by environmental stress or pest invasion, called induced secondary compounds (opens in new window). The latter requires a higher energy cost to the plant.
Some plants, mainly those found in the legume family (Leguminosae), can utilize nitrogen (N2) from the atmosphere by converting it to a useable form (ammonium, NH4+) with the help of a rhizobia bacteria. In this symbiotic relationship, the bacteria invade the roots and create nodules where they fix nitrogen into a usable form for plant use. This is called nitrogen fixation (opens in new window). Most plants utilize nitrogen from the soil that has already been transformed into the ammonium and nitrate (NO2–) by soil fungi and bacteria, a process called nitrification.
Many root systems work together with soil fungus in a mutually beneficial relationship. This root-fungus association is called mycorrhiza (opens in new window), literally meaning fungal root. The plant provides food made through photosynthesis to the fungus, and the fungus helps supply water and nutrients to the plant as root extensions.
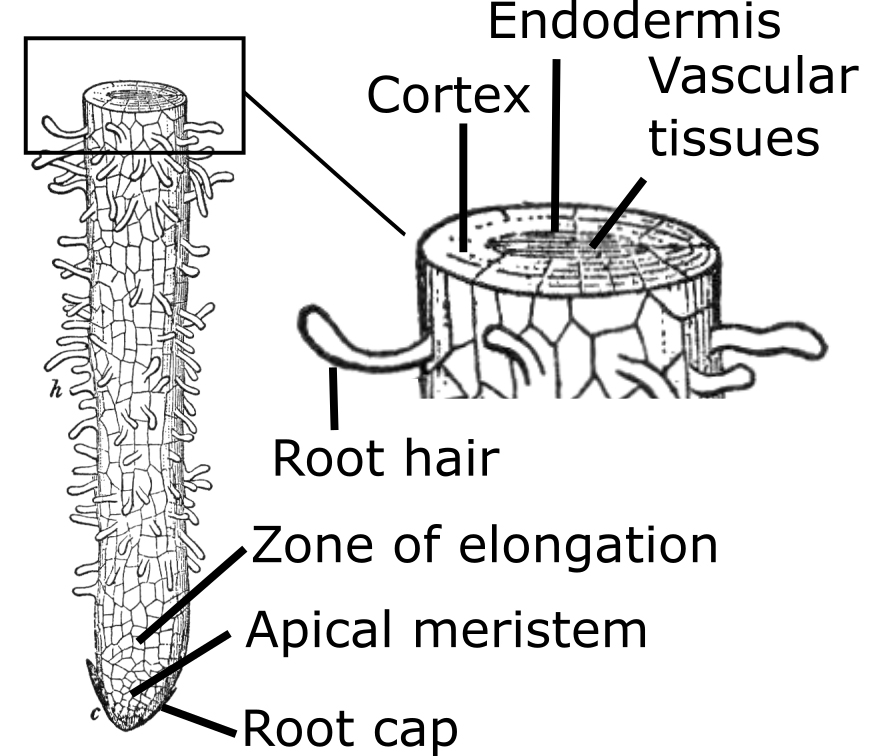
Root Structure. Roots have an epidermis layer with root hairs that are single-celled extensions of epidermal cells (Figure 6). Root hairs provide the water and nutrient uptake for a plant. They are delicate structures that are easily killed by desiccation or during transplanting. Roots typically do not have buds or leaves.
Interior tissues of roots include the cortex, vascular tissues, and the endodermis. The cortex is responsible for water and mineral transport and storage; the vascular tissues transport water, nutrients, and other compounds produced in the root upward and food and other compounds down from the top of the plant; and the endodermis helps control water flow and is the tissue from which side or branch roots are generated.
Root Growth. Root growth is very similar to that of a stem in that there is an apical meristem at the root tip. Behind the apical meristem there is the zone of elongation, where the newly formed cells stretch out and push the tip further into the soil. Behind the zone of elongation there is the zone of differentiation where the unspecialized elongated cells develop into the discrete tissues of bark, phloem, cambium, xylem, cortex, and endodermis tissues.
There are many differences between a stem and a root. For instance, the growing root tip is covered with a hard layer of cells called the root cap which is produced by the apical meristem. The root cap is a sort of battering ram that forces its way between, and sometimes through, soil particles. The pressure that forces the root cap through the soil comes from the expansion of newly formed cells in the root apical meristem and the expanding cells in the zone of elongation. Behind the apical meristem, in the elongation zone, is where root hairs are produced. Root hairs are very short-lived, from a few hours to a few days. They are the primary water- and nutrient-absorbing structure for the root and, therefore, the plant.
Types of Roots. Roots may simply be classified as fibrous, tap, or fleshy. Fibrous roots (opens in new window) are generally considered to be stretched out flat and somewhat close to the soil surface, while taproots (opens in new window) are usually depicted as growing straight downward from the surface. Fleshy roots may be configured as either taproots or fibrous roots.
To a certain extent, root configuration is genetically controlled. Under good conditions, planted in the ground with a well aerated, deep, moist soil, maples (Acer spp.) will usually have a rather flat, shallow, fibrous root system, while trees like honey locust (Gleditsia sp.) and redbud (Cercis sp.) will tend to have a more spreading, penetrating root system. Hickories (Carya spp.) and walnuts (Juglans spp.) are considered to be taprooted. Food plants such as radishes, carrots, beets, turnips, and rutabagas are also examples of not only taproot systems but fleshy roots. Other forms of fleshy or tuberous roots are dahlias (Dahlia spp.) and sweet potato/yams (Ipomea batatas). Plants growing in containers may not develop the normal root configuration characteristic of the species due to soil volume and pot edge constraints.
Adventitious roots are any roots arising at an unexpected place on a plant. The short, white bumps along a tomato stem are adventitious roots. Aerial roots are adventitious roots that develop from aboveground stem tissues. Aerial roots may grow to support the plant, as with English ivy (Hedera spp.). Aerial roots are common with plants such as pothos (Epipremnum aureum) and philodendron (Philodendron spp.).
Root Extent and Depth. There are many misconceptions and misinformation about the extent and depth of plant roots in both natural and human-dominated landscape settings. Roots basically need two things: gaseous exchange for respiration and soil water with its dissolved mineral nutrients. If either air or water is lacking, the roots will not grow. In attempts at plant salvage in the arid regions of the southwestern United States, workers very often find that plant roots extend 12 to 14 feet or more below the surface, where water is present. This condition is not normally found in highly compacted, human-dominated landscape soils.
Where the soil is saturated with water, air is mostly excluded—as is the oxygen in the soil air. Respiratory carbon dioxide cannot be eliminated and therefore accumulates in root tissue with serious side effects. In this case the roots generally grow more toward the surface and sometimes on top of the surface. Common causes for soil saturation include a high water table; a hard, impenetrable layer somewhere below the surface with a secondary water table above it; very tightly compacted, poorly drained soil, a common occurrence in many developed residential areas; and just plain overwatering, especially on poorly drained soils.
Leaves
There are many different types of plant leaves. The “typical” leaf is a flat, blade-like structure, attached to the stem (usually) by a stalk called a petiole (opens in new window). The juncture of the leaf and the stem is called the leaf axil.
Leaf Function. Leaves are where the majority of the work of a plant occurs. Leaves are the main site of photosynthesis, respiration, transpiration (opens in new window), and hormone synthesis. Leaves also move water, mineral nutrients, and materials made by the roots around in the leaf and move manufactured food and other metabolites out of the leaf (more on these processes later in the chapter). Leaves provide storage of food and sometimes water, and synthesize hormones that are used throughout the plant.

Leaf Structure. Leaves of dicot plants have net venation, also called reticulate venation, as seen in the image on the left in Figure 7, while leaves of monocot plants typically have parallel veining as seen in the image on the right in Figure 7. Veining patterns are helpful in plant identification and are discussed in detail in Chapter 2: Plant Nomenclature and Identification.
Conifers may have needle-like leaves (Douglas-fir, Pseudotsuga menziesii; hemlock, Tsuga spp.; ponderosa pine, Pinus ponderosa), scale-like leaves tightly pressed onto the stem (arborvitae, Thuja spp.; California incense cedar, Calocedrus decurrens) or awl-like leaves, which are short, narrow, sharp-pointed leaves, usually pulled away from the stem (junipers, Juniperus spp.; giant redwood, Sequoiadendron giganteum) (Figure 8). Junipers may have two or three different types of leaves on the same plant.

A “typical” leaf has four layers of different types of tissue (Figure 9). The top layer is the upper epidermis, and its primary function is protection. The epidermis may be covered with a waxy cuticle, or various types of hairs, waxy scales, or other forms of protective structures. The next layer contains palisade cells (opens in new window), which are elongated cells set perpendicular to the upper epidermis. These cells are chlorophyll-rich and this is one of the areas where photosynthesis takes place. Underneath the palisade layer is the spongy mesophyll (opens in new window) layer which not only has photosynthetic capabilities but has voids between the cells to facilitate gaseous exchange. The bottom layer is the lower epidermis which provides protection.

The vascular bundles with xylem and phloem run through the spongy mesophyll, transporting water, nutrients, and photosynthate (the products of photosynthesis) into and out of the leaf tissue.
The lower epidermis has openings called stomata (opens in new window) (singular: stoma) that allow gases to enter and exit the leaf (some plants also have some stoma on the upper surface). Each stoma has two guard cells that open and close to allow oxygen, water vapor, and other gases to exit and air (with carbon dioxide) to enter. They are closed in the absence of light and when the plant is under water stress. This limits the amount of water the plant loses during dry periods but also limits the amount of carbon dioxide taken in and, therefore, reduces the photosynthetic activity.
Leaf abscission is the process whereby a leaf falls off the stem. A layer of cells between the leaf and the stem, known as the abscission layer (opens in new window), is killed by plant hormones at the end of the growing season. On deciduous plants, this process occurs annually. On evergreen plants it takes two or more years. Thus, evergreen plants, while always green, do lose their leaves—just not all at once to go bare like deciduous trees.
Leaf Modifications. As with stems, leaves may be specially modified to provide a particular function for a plant. Leaves may be modified to provide nutrient storage (onion, Allium spp.) or water storage as in stonecrops (Sedum spp.) and hens-and-chicks (Sempervivum spp.).
Cotyledons are the seed leaves of a germinating dicot seed. They usually do not resemble the true leaves. They may come out of the ground on germination, epigeous germination (opens in new window) (cabbage, tomato), or stay underground, hypogeous germination (opens in new window) (oaks, Quercus spp.).
Flower bracts (opens in new window) are modified leaves found just below a flower. They can be smaller than petals but may also be larger and of colors other than green. For example, the white showy part of the flowering dogwood (Cornus florida) and the (usually red) showy part of the poinsettia flower are, in fact, bracts (opens in new window). In both of these cases, the actual flower is less noticeable than the bracts and yellow in color. Another example of bracts is the whorled short green “leaves” found at the base of a sunflower.
Bud scales, also called cataphylls, are modified leaves that cover the buds of woody plants. On a cactus plant (Echinocereus spp.), spines are leaves modified for protection, whereas thorns on hawthorn (Crataegus spp.) are modified stem tissue, and prickles on roses (Rosa spp.) are outgrowths of the stem cortex and dermal tissue. In some species such as grape (Vitis spp.) and clematis (Clematis spp.), tendrils may help plants climb and are usually modified leaf tissue, but sometimes modified stem tissue.
Flowers
Flowers are usually the showy part of a plant. Many gardeners grow plants specifically for their flowers. But flowers serve other useful purposes: they are the reproductive organs a plant uses to form seeds that reproduce its species, and they are the basis for the classification of flowering plants.
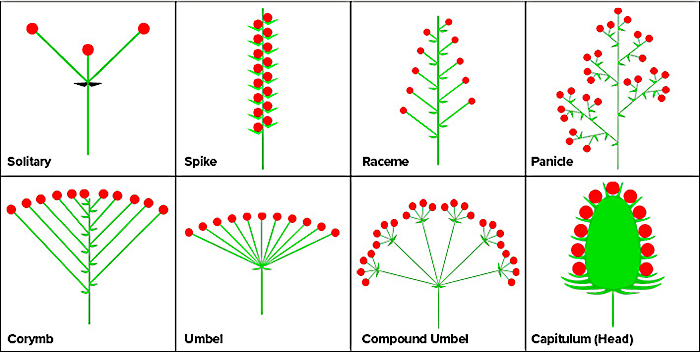
The so-called “flower” of a plant, such as lilac (Syringa vulgaris), daisy (Leucanthemum spp.), or hydrangea (Hydrangea spp.), is actually an inflorescence, which is a group of individual flowers arranged in a cluster. Inflorescences (opens in new window) can be found in many different shapes and sizes, and they can be arranged on a plant in a number of ways. A wide variety of inflorescence occur in nature. They can be simple (one flower on a stalk) or compound (multiple flowers on a stalk). A few common types are solitary, spike, raceme, umbel, corymb, capitulum, and panicle, as seen in Figure 10.
Common Inflorescence Types
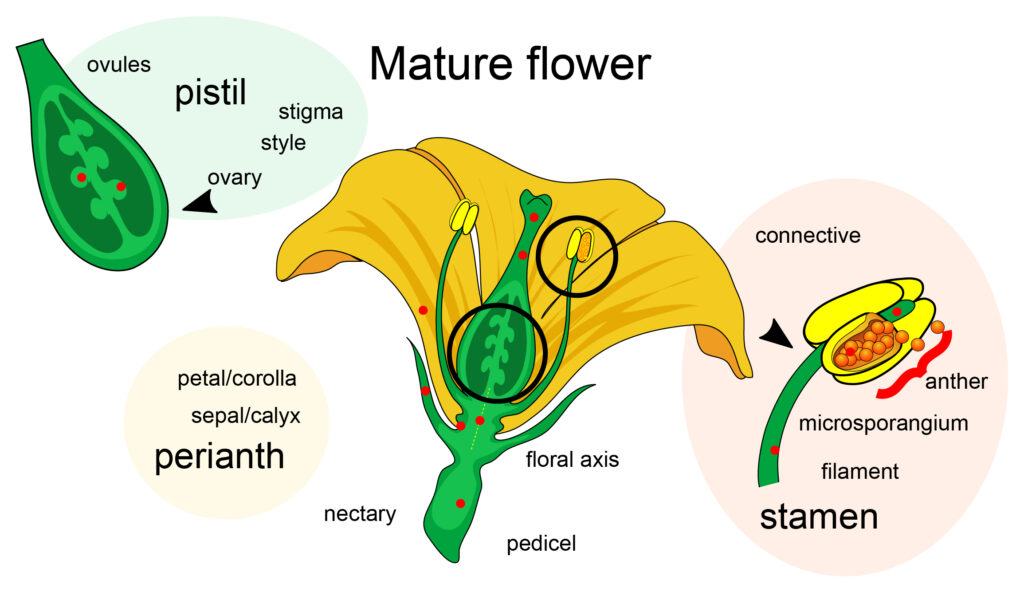
Flower Parts. Sepals (opens in new window) are modified leaves which enclose and protect the flower bud. Collectively, they are called the calyx. The sepals may fall off after the flower opens, remain attached under the petals, or may expand and be very prominent and look similar to the petals. Petals are the typical showy part of the flower; together, the petals of the flower make up the corolla (opens in new window). The calyx and the corolla are vegetative, non-reproductive parts of the flower collectively called the perianth.
There are many modifications of flowers. For instance, petals, sepals, or both may be fused together to form a tubular flower, as with penstemon (Penstemon spp.).
Stamens (opens in new window)are the male part of the flower, consisting of the filament (opens in new window), a stalk which holds up the anther (opens in new window). The anther is the structure which contains the pollen, and the pollen contains the male sex cells.
The female portion of the flower is the pistil (opens in new window). It consists of a sticky surface called the stigma which is held above the ovary by the style. The ovary contains the ovule (egg cell or cells) which, when fertilized, will become the embryo of the seed. The ovary will develop into other parts of the seed and the fruit typical of the species.
The pistil and stamen are the reproductive flower parts. A perfect flower is a term for a flower that contains both male and female parts. A complete flower contains male and female parts plus petals and sepals (Figure 11). A flower is considered imperfect if it contains only male or female parts. Pistillate flowers have only female parts (no male parts are present), and staminate flowers have only male parts (no female parts present).
Monoecious (meaning “of one house”) plants have separate male and female flowers on the same plant, while dioecious (meaning “of two houses”) plants have male flowers on one plant and female flowers on another.
Pollination (opens in new window) is the process of a mature pollen grain from the anther landing on the mature stigmatic surface of the pistil. The agents for pollination are usually insects or wind, but there are others. The pollen grain “germinates” on the receptive stigmatic surface and a pollen tube containing the male sex cells grows down the style to the ovule or egg cell. The union of the male and female cells is called fertilization. The fruit will then develop in the form that is typical for the species. It is important to note that if the pollination–fertilization process does not occur, normally no fruit will form.
Self-pollination (opens in new window) is the transfer of pollen from the anther to the stigma of the same plant or the same clone. Cross-pollination (opens in new window) is the transfer of pollen from one clone to another clone or from one flowering plant to another flowering plant. Cross-pollination is necessary for some plants to bear fruit. The apple cultivars ‘Liberty’ and ‘Chehalis’ are not self-fruitful (opens in new window), but ‘Liberty’ can cross-pollinate ‘Chehalis’ and vice versa. In order for two plants to cross-pollinate, they have to be in bloom at the same time, and the agent of transfer (bees, wind, etc.) has to be present and active.
Buds (opens in new window). Buds are undeveloped new shoots. Active buds are those buds that will do something during the next or current growing season. They will become leaves, shoots, flowers, or a combination of leaves and flowers. Latent buds (sometimes called epicormal buds) are formed in the normal manner, but do not grow and develop without special stimulus. These buds will remain dormant and under the bark until they are forced to grow, either by a pruning cut or limb breakage. This is one of the reasons why, when a large limb is broken off or an improper stub cut is made, shoots are formed below the injury. This is a survival mechanism that plants have.
Adventitious buds (opens in new window) are those buds which form in a position on the plant where they are not normally found. This includes bud formation on the internode portions of the stem. This sometimes comes about by radical limb removal (stub cuts on large limbs) or limb breakage because of severe winter weather. The important difference between latent buds and adventitious buds is that the latent buds are formed in the normal manner, and adventitious buds on stems are usually formed in response to injuries, either pruning or limb breakage.
Flowering Patterns. It is important to know when a plant sets up, or initiates, its flower buds and where those buds are, especially if pruning is to be done to enhance flowering. There are a number of different blooming patterns of woody plants.
Woody plants may bloom from buds that were initiated the previous fall, prior to the spring in which they bloom. They may bloom on the terminal end of the branch from simple buds (star magnolia, Magnolia stellata; Burkwood viburnum, Viburnum ×burkwoodii). They may also bloom from the end of the stem from a mixed bud (horsechestnut, Aesculus hippocastanum; Norway maple, Acer platanoides). Some woody plants bloom on the end of short, specialized branches, called spurs (opens in new window), that have very short, compressed internodes (apples, Malus spp.; chokeberries, Aronia spp.; serviceberries, Amelanchier spp.).
Woody plants can also bloom from the lateral portions of the stem which was produced the previous year. These may be from a simple bud (forsythia, Forsythia sp.; Thunberg’s spirea, Spiraea thunbergii; bridal wreath spirea, S. prunifolia) or a mixed bud (large-leaf hydrangea, Hydrangea ×macrophylla; red currant, Ribes sanquineum; Vanhoutte’s spirea, Spiraea ×vanhouttei; most mock oranges, Philadelphus spp.; and weigelas, Weigela sp.).
Many later-blooming shrubs and trees set up their flower buds as they are growing. These include PeeGee hydrangea (Hydrangea paniculata grandiflora), Bumalda spirea (Spiraea × bumalda), and Blue Mist (Caryopteris clandonensis). Note that both spirea and hydrangea include some species that initiate their flower buds in the fall and other species which set up flower buds as they are growing.
Flowering and Fruiting. A plant must flower before it can bear fruit (or set seed). Flower buds on many species of temperate zone plants are formed in the late summer or early fall prior to the spring in which they bloom. These plants usually bloom in early to late spring. Other plants set up their flower buds after they break dormancy in the spring of the year. These plants generally bloom later in the summer and into early fall.
Failure to fruit could result from a number of causes:
- Lack of pollination; absence of bees or other pollinators; too cold or wet for the bees to fly.
- Plant didn’t flower that year; plant is too young to flower.
- Flower buds were frozen.
- Flower buds were pruned away.
- Plant only has male flowers (as with Ginkgo biloba or holly, Ilex sp.).
- Lack of a cross-pollinator or the cross-pollinator blooms at a different time.
- Alternate-year blooming occurs: having a heavy crop one year leaves a lack of energy to produce many flower buds for the following year.
As with flowers, fruits come in many different sizes and shapes, as seen in Figure 12. They are classified based on factors such as number of seeds per fruit, segmentation, fleshiness, seed placement within the fruit, and other characteristics. A few examples of fruit types include:
- Simple—pome (apple, Malus spp.; mountain ash, Sorbus spp.); drupe (cherry, Prunus spp.; viburnum, Viburnum spp.); berry (tomato, Solanum lycopersicum).
- Aggregate—(strawberry, Fragaria spp.; raspberry, Rubus spp.).
- Dry—capsules (mock orange, Philadelphus spp.; rhododendron, Rhododendron spp.), samara (maple, Acer spp.), nuts (walnut, Juglans spp.), pod (goldenchain tree, Laburnum spp.; honeylocust, Gleditisia spp.).

Plant Growth and Development
Metabolism
Metabolism is the sum total of the biochemical activity within a living organism. Some of the chemical reactions take energy to complete, others release energy when completed.
Photosynthesis
Photosynthesis is an energy harnessing process. Simple carbohydrates (glucose) are formed by combining carbon dioxide (CO2) and water (H2O), using the radiant energy from the sun and the catalyst, chlorophyll. Oxygen (O2) is given off as a byproduct. During this process the radiant energy from the sun is converted to chemical energy and is stored in the chemical bonds of the sugar molecule that was formed.
Carbon dioxide comes into a leaf via the stomata with the air, while the water and other necessary nutrients are absorbed by the roots.
Some of the factors affecting the rate of photosynthesis include:
- The amount of incoming solar radiation.
- Quantity of chlorophyll-containing cells: leaves developed in the full sun usually have less chlorophyll-bearing cells by leaf area weight than shade leaves.
- Plant and leaf age: older plants and older leaves have a lower photosynthetic rate than younger plants and leaves do.
- Carbon dioxide levels: increasing carbon dioxide levels above the normal atmospheric 400 ppm can increase the rate of photosynthesis, up to a point. Some growers of greenhouse crops will enrich the greenhouse air with carbon dioxide by burning propane. This is called carbon dioxide fertilization (opens in new window).
- Air and soil temperatures: warmer temperatures increase the rate of photosynthesis, up to a point. The optimum temperature for photosynthesis is 95–104°F. Higher temperatures slow it down as the enzymes needed for the reaction start to break down.
Chlorophyll contains carbon, oxygen, hydrogen, nitrogen, and magnesium. Iron (Fe) and manganese (Mn) are two other elements essential for the synthesis of the chlorophyll. Carbon, oxygen, and hydrogen are normally not lacking in plants. If iron, manganese, or magnesium (Mg) becomes deficient in a plant, there will be less chlorophyll manufactured, and the leaves will look more yellow (chlorotic (opens in new window)) because there is less chlorophyll.
Respiration
While photosynthesis is an energy-harnessing process, respiration is an energy-releasing process where carbohydrates (usually, but other metabolites may be used) are combined with oxygen (through oxidation) resulting in the release of energy, and with carbon dioxide and water given off in the process. The energy liberated by this process may be used to drive other chemical reactions such as the synthesis of proteins or the many other materials the plant needs.
Respiration is photosynthesis in reverse. The energy delivered originally by the sun is now converted to energy the plant can use in chemical reactions and in other processes. Photosynthesis only occurs in the light; respiration occurs in both light and dark. The rate of respiration increases with increasing temperatures.
To recap:
Photosynthesis:
Carbon dioxide + water + (radiant) energy + catalyst (chlorophyll) => glucose + oxygen
Respiration:
Glucose + oxygen => carbon dioxide + water + (chemical) energy
Transpiration
Water vapor escapes a leaf through the stomata when their guard cells are open. This process is called transpiration. Water movement into the roots, up the stem, into the petiole, and out the leaf stomata is called the transpiration stream. The guard cells, surrounding each stoma, are like little balloons. When they are filled with water, they bend to open, and water vapor (and oxygen from photosynthesis) can escape while air with its carbon dioxide can enter. When guard cells lose water they become soft and close, keeping the water and oxygen in and air (and carbon dioxide) out. Stomata are only open under light conditions (natural or artificial) and when the plant is under good water relations. In dark or dry, droughty situations they will close, minimizing water loss. Under dark conditions transpiration stops and so does the photosynthetic process.
Plant Hormones
Plant hormones are chemicals produced by the plant in small quantities. Hormones are transported to specific sites within the plant where they have a pronounced effect on some aspect of plant growth and development. The main hormones important for plant growth and development are listed in the Examples of Common Plant Hormones sidebar.
Plants have the ability to synthesize hormones and they also have the ability to stop the action of those hormones when necessary by breaking them down or by inhibiting or slowing down their synthesis. The synthesis and inhibition of plant hormones is regulated by many internal and external influences including temperature, drought, and plant defense.
Aspects of Light
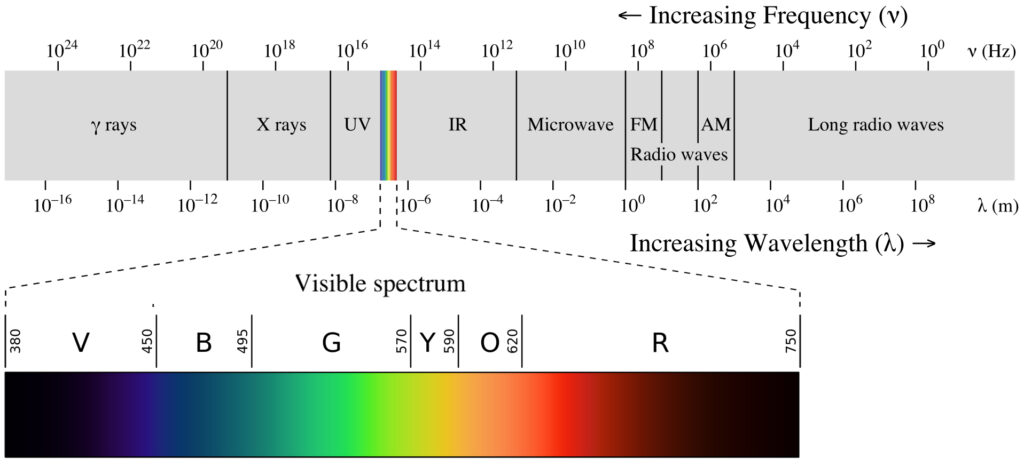
Light is a form of electromagnetic energy released as photons. The terms quantity, duration, and quality are used when describing light.
Light quantity relates to the intensity or the energy output of a light source.
Light duration (opens in new window) indicates the photoperiod or length of time that the light source is emitting light.
Light quality (opens in new window) refers to the color of the light emitted by a source. A light’s color is determined by the light’s wavelength (measured in nanometers, nm). Sunlight is energy of wavelengths from the entire electromagnetic (EM) spectrum. The wavelengths of light that humans can see make up the visible spectrum (opens in new window)—just a small portion of the EM spectrum, from violet to red light. Wavelengths within the visible spectrum are most effective in stimulating photosynthesis and are referred to as the photosynthetically active radiation (opens in new window) (PAR).
Objects appear colored because of reflection: they reflect light of a certain wavelength. Green plant leaves appear green to our eyes because they reflect green wavelengths while absorbing the other wavelengths of light. A black object absorbs all wavelengths of light and reflects almost none, while a white object absorbs almost no light and reflects nearly all wavelengths.
Light quantity, or intensity, is governed by time of year and day, cloud cover, and shade from overhead vegetation, buildings, and anything casting a shadow. Shade can reduce the growth rate of plants because the lower light level decreases the rate of photosynthesis.
The light compensation point (opens in new window)is the level of light intensity at which the rate of photosynthesis (and a plant’s CO2 requirement) equals the rate of respiration (the plant’s CO2 output).
Photoperiodism (opens in new window) is the induction of some change in a plant’s growth and development by a change in the relative length of light and dark periods (i.e., night and day). The best examples of this are the initiation of flower bud formation and onset of bud dormancy (opens in new window) in the later summer and early fall period.
Flower bud formation is governed by a critical day length. A long-day plant (opens in new window) (actually, a short-night plant) will set up its flower buds if the critical day length is exceeded (days getting longer). If a plant has a critical day length of 11.6 hours, this length of light must be exceeded before the plant will initiate flower buds. For short-day plants (opens in new window) the photoperiod must be less than the critical photoperiod (days getting shorter). Typically, short-day plants bloom more toward the end of summer (chrysanthemums, Chrysanthemum spp.) and long-day plants flower more toward the spring–early summer period (lilac, Syringa spp.).
Day-neutral (opens in new window) plants are not controlled by any specific photoperiod and bloom when they are mature enough to flower and the environmental conditions are suitable for that species to flower (corn, Zea mays; dandelion, Taraxacum spp.; rose, Rosa spp.; and grapes, Vitis spp.).
The shorter days of late summer and early fall initiate the onset of bud dormancy. Hormones chemically lock up the buds to prevent buds from growing in the fall and during mild spells in the winter. Also, during this period of shortening days, other metabolites are synthesized by the leaves and are translocated to other parts of the plant, slightly increasing the hardiness of the plant parts. To overcome this physiological, chemically induced dormancy, the plant usually needs a cold or chilling period for a certain length of time. During this chill period the concentrations of growth inhibiting substances (such as ABA) in the plants are diminished and the concentrations of growth promoting substances (such as GAs) increase.
Temperature
Temperatures are governed by a number of different factors and have many different and pronounced effects on plant growth. Incoming solar radiation, which heats land masses, bodies of water, plants, buildings, etc., is influenced by cloud cover, air pollution, and day length (season). Bodies of water (large rivers, lakes, bays, oceans) heat and cool slowly in comparison to land masses which tend to heat and cool rather rapidly. Even wet, heavy soils will warm up much more slowly than lighter (sandy) soils that do not contain much water.
Cold air is more dense than warm air and will sink to low areas in a landscape. When cold air is regularly trapped in a particular low area, that area is called a frost pocket (opens in new window).
Other temperature considerations:
- Increasing temperatures will increase the rates of photosynthesis, respiration, and transpiration.
- A certain period of time at chilling temperatures (40–45°F, or 4–7°C) is necessary for overcoming physiological dormancy of certain plants. The length of chill needed will vary with the species and even, in some cases, with cultivars or ecotypes within a species.
- There are combinations of light and temperature such as long days and warm temperatures to initiate flowering. Southern magnolia (Magnolia grandiflora) and crapemyrtles (Lagerstroemia indica) are examples. Flowering of both of these species may be diminished by cooler summer temperatures.
- Certain vegetable crops need warmer temperatures to develop or develop properly, while others do much better with lower summer temperatures. Examples of warm season crops are peppers (Capsicum spp.) and melons (Citrullus spp. and Cucumis spp.), while cool season crops would include examples such as lettuce (Lactuca spp.) and cole crops (Brassica spp.). For more discussion, see Chapter 7: Vegetable Gardening.
Higher temperatures may cause some plants to bolt (opens in new window) or start to flower. Examples are cilantro (Coriandrum sp.) and lettuce.
- Some seeds need a chill period to germinate, called stratification. Examples are larkspur (Consolida spp.) and lupine (Lupinus spp.).
- Seeds may germinate over a wide range of temperatures, but most plants have an optimum germination temperature at which the seeds will germinate more quickly and uniformly.
- Tolerance of a plant and its parts to both cold and heat is called its cold hardiness. For further discussion of plant hardiness, see below and Chapter 11: Herbaceous Landscape Plants.
- Vernalization is a chilling requirement that some plants need to initiate flower buds. Examples of this are tulips (Tulipa spp.) and winter wheat (Triticum spp.). Tulips are planted in the fall to get a winter’s chill period in order to form a flower bud for spring blooming.
Water
Water has many functions and roles in plant growth and development. Water is a solvent for the materials entering and moving within the plant, and it is a reactant in many of the internal chemical reactions going on inside the plant. For instance, water is a major constituent of photosynthesis. Water also regulates the turgor pressure (turgidity) of cells, thereby affecting transpiration and cell growth. Similarly, it regulates stomatal opening and closing, thereby regulating transpiration and photosynthesis within a plant. And finally, water is a major agent of evaporative cooling of leaves—transpiration.
Relative humidity (opens in new window) (RH) is the amount of water vapor found in the air compared to the amount of water vapor that the air can hold at that particular temperature, expressed as a percent. Air can hold more water vapor at a higher temperature than it can at a lower temperature. Water vapor will move from an area of high RH to an area of low RH. The RH inside a leaf is very high, so therefore water vapor has a tendency to move out of the leaf, to an area of lower RH. There are RH applications in plant propagation where mist and fogging systems have been developed to keep the RH elevated around cuttings (reducing transpiration) while the cutting develops new roots.
Nutrition
Nutrients have specific roles in plant growth. Nutrients may be required as part of some molecular structures or they may be necessary as a sort of catalyst to help complete some biochemical reactions.
Nutrients have to be soluble in water to enter a plant, and they have to be soluble to move within the plant. A nutrient’s concentration in plant tissue is usually expressed in parts per million (ppm).
Nutritional elements may also be classified as mobile (opens in new window) or non-mobile (opens in new window). Mobile elements will move in the plant toward the growing point or toward younger leaves. Non-mobile elements will usually stay put once they have entered the leaf. When a mobile nutrient element is lacking, that nutrient is moved to new growth areas, so the deficiency symptom will show up in the older leaves. Conversely, the lack of a non-mobile element will become apparent in the newer leaves since the nutrient cannot move to the growing points.
See Chapter 4: Plant Mineral Nutrition and Fertilizers for an in-depth discussion of nutrients and how plants use them.
Plant Hardiness
Plant hardiness may be defined as a plant’s tolerance to extremes. The extremes could be to temperature, water availability, soil pH, etc. In most cases though, hardiness refers to the cold hardiness of the plant.
Temperate zone plants develop hardiness in a predictable manner. The initial stages of cold hardiness and physiological dormancy development begins with shortening days in the late summer, early fall. This process can increase the hardiness of the plant up to about 10°F. Plants at this stage can endure the early fall frosts with little or no freeze damage.
As temperatures start cooling in the fall, plants adapt by acclimating, or hardening, to cold. This process continues until the plant reaches its ultimate mid-winter hardiness, which is genetically controlled. African violets can be killed by chill damage at temperatures slightly below a mere 40°F. Arborvitae (Thuja spp.) can develop cold hardiness down to −100°F.
Plants will de-acclimate, or become less hardy, with increasing temperatures in the latter part of winter and the very early spring. Most plants, when they start to grow or are in active growth, have very limited or no tolerance to freezing temperatures.
Freeze damage, usually not apparent until the following spring, can occur in a variety of situations. All of the below have occurred in Washington on numerous occasions in the last 20 years. Some occur on a yearly basis.
- Very warm fall days (Oct./Nov.) may provide no impetus for hardening or acclimating to cold, followed by a dramatic and sudden drop in temperature well below freezing. It seems this usually happens in late fall.
- Temperatures drop below the (genetic) ultimate mid-winter hardiness of the plant.
- A week to 10 days of abnormally warm temperatures in mid to late winter can lead plants to de-acclimate. That balmy period will be followed by a dramatic and sudden drop in temperature back to normal below-freezing temperatures. Similarly, this may also happen when evergreen foliage or tree bark heats up on the southwest side of the plant on a cold (below freezing), sunny, winter day. The plant leaves or bark can reach temperatures of 50°F or more from the impact of solar radiation. When the sun goes down, the plant tissue very rapidly, often within minutes, cools down to the ambient air temperature which is below freezing. Cellular water that was loosened up during the heat of the day cannot be withdrawn quickly enough during the very rapid cooling process, so ice crystals form in the cells, bursting them and resulting in tissue death.
- Spring frosts after plants have begun to grow can injure any new growth. When plants start to grow in the spring, the new tissues have very limited or no hardiness. Late freezes affect not only leaves and shoots but also flowers and early setting fruit such as apricot. The flowers of star magnolia (Magnolia stellata), saucer magnolias (M. soulangiana), and Higan cherry cultivars (Prunus subhirtella) are very susceptible to any type of freeze when they are in bloom. In areas where late frosts happen very frequently, substitutions of other, later blooming species, or cultivated varieties of the magnolias and cherries mentioned above may be made.
Ultimate mid-winter hardiness of plant parts varies as follows: stem tissue is usually hardier than vegetative buds, which are usually hardier than flower buds, which are usually hardier than roots. The bottom line is that, in some cases, stems may be hardier by 30 to 40°F (or more) than the roots.
Zones of cold hardiness have been mapped by the USDA (Figure 13).

Learning More
Many good resources on various aspects of botany and plant sciences exist. They vary in their scope of coverage on different topics and in the depth of coverage, ranging from basic gardening books with a single chapter on botany to in-depth college-level texts. Reference materials on specific subjects within botany such as plant anatomy, plant physiology, plant geography, taxonomy, plant ecology, and more can be found online, on gardening apps, and in printed material. Many universities have excellent resources for download.
A good general horticulture book usually has chapters on botanical principles as related to the various horticultural crops. These textbooks and references are usually a little bit more practically oriented.
Further Reading
WSU Extension publications (opens in new window).
Ambroise, V., S. Legay, G. Guerriero, J.-F. Hausman, A. Cuypers, and K. Sergeant. 2020. The Roots of Plant Frost Hardiness and Tolerance (opens in new window). Plant and Cell Physiology 61(1):3–20.
Bielczynski, L.W., M.K. Łącki, I. Hoefnagels, A. Gambin, and R. Croce. 2017. Leaf and Plant Age Affects Photosynthetic Performance and Photoprotective Capacity. Plant Physiology 175(4):1634–1648.
Bienz, D.R. 1993. The Why and How of Home Horticulture. San Francisco: W.H. Freeman and Co.
Buis, A. 2019. The Atmosphere: Getting a Handle on Carbon Dioxide (opens in new window). NASA Jet Propulsion Laboratory. NASA Global Climate Change, Vital Signs of the Planet.
Brickel, C., and J.D. Zuk, eds. 2004. The American Horticultural Society A–Z Encyclopedia of Garden Plants, revised edition. New York: DK Publishing.
Capon, B. 2010. Botany for Gardeners, 3rd edition. Portland, OR: Timber Press.
Chalker-Scott, L. 2015. How Plants Work—The Science Behind the Amazing Things Plants Do. Timber Press.
Elpel, T.J. 2013. Botany in a Day, The Patterns Method of Plant Identification. Hops Press LLC.
Evert, R.F., and S.E. Eichhorn. 2013. Biology of Plants. W.H. Freeman and Company Publishers.
Greene, D.W., and W.R. Autio. 1994. Notching Techniques Increase Branching of Young Apple Trees (opens in new window). Journal American Society Horticulture Science 119(4):678–682.
Liu, L., N. Lin, X. Liu, S. Yang, W. Wang, and X. Wan. 2020. From Chloroplast Biogenesis to Chlorophyll Accumulation: The Interplay of Light and Hormones on Gene Expression in Camellia sinensis cv. Shuchazao Leaves. Frontiers in Plant Science 11:256. DOI: 10.3389/fpls.2020.00256.
Mabberley, D.J. 2008. Mabberley’s Plant-Book, 4th edition. New York: Cambridge University Press.
Martin, R.E., G.P. Asner, L.P. Bentley, A. Shenkin, N. Salinas, K.Q. Huaypar, M.M. Pillco, et al. 2020. Covariance of Sun and Shade Leaf Traits Along a Tropical Forest Elevation Gradient. Frontiers in Plant Science 10:1810. DOI: 10.3389/fpls.2019.01810.
McCree, K.J. 1981. Photosynthetically Active Radiation (opens in new window). In Physiological Plant Ecology I. Encyclopedia of Plant Physiology, O.L. Lange, P.S. Nobel, C.B. Osmond, and H. Ziegler, eds., (New Series), vol 12 / A. Springer, Berlin, Heidelberg.
National Agricultural Library (opens in new window). 2010. United States Department of Agriculture.
Runkle, E. 2015. Interactions of Light, CO2 and Temperature on Photosynthesis (link to PDF document). Michigan State University Extension.
Stark, G. 2020. Light (opens in new window). Encyclopedia Britannica.
Stearn, W.T. 2004. Botanical Latin. Portland, OR: Timber Press, Inc.
USDA ARS (opens in new window) (Agricultural Research Service). 2023. Hardiness Zone Map.
USDA NRCS (opens in new window) (Natural Resource Conservation Service). n.d. PLANTS Database.
Whiting, D., A. O’Connor, H. Hodgin, J. Jones, L. McMulkin, and L. Potts. 2009. Taxonomic Classification (link to PDF document). Colorado State University Extension Publication CMG Garden Notes #122.
Yingying, H., H. Yang, and Y. Jiao. 2014. Regulation of Inflorescence Architecture by Cytokinins (opens in new window). Frontiers in Plant Science 5.